


On scientific laws, classic dances, and a new study about molecular evolution…in reverse.
Guest post by C. Brandon Ogbunu
Taking direction from evolution
One feature of evolutionary biology that has always intrigued me is the preponderance of “laws.” That is, not “laws” as in Newton’s laws of motion, but less formal ones—analogies and principles that describe a theoretical idea. Many do not have the term “law” attached to them but are laws nonetheless: terms and phrases that describe presumptive fundamental evolutionary phenomena, like the Red Queen hypothesis, Muller’s ratchet, and punctuated equilibrium.
I have a longstanding curiosity about the origins and role of these laws in biology; I think some of them stick around because they help communicate complicated ideas, even if they’re arbitrarily constructed or imprecise.
Dollo’s law is one of my favorite examples:
“An organism is unable to return, even partially, to a previous stage already realized in the ranks of its ancestors.”
In the Blind Watchmaker, Richard Dawkins interpreted it as:
“….just a statement about the statistical improbability of following exactly the same evolutionary trajectory twice (or, indeed, any particular trajectory), in either direction.”
Or even more casually (as I say it):
“It is difficult for an improbable event to happen and then un-happen in the exact same way.”
Essentially, Dollo’s law speaks to the improbability of “reverse evolution.”
Now before you jump in, I know, I know—there are no real directions in evolution. So what am I getting at?
Sure, there is no true “forward” or “backwards” in evolution, and the general misuse of the language of progress has been costly to evolutionary biology. Too many of us (even practicing biologists) tend to erroneously discuss or think about evolution as being a progressive force.
That said, we can discuss direction when we are talking about the evolution of very specific traits in specific settings. For example, if we talk about a population of bacteria evolving resistance to an antibiotic, we can talk about evolving towards resistance (for conversation purposes, at least). “Reverse,” in this situation, would be from a resistant form “back” to a treatable form.
Why is this important?
Well, one of the proposed strategies for preventing the evolution of antibiotic resistance is to drive evolution “backwards.” If a population is resistant to available antibiotics, perhaps we can use drugs to encourage that population to evolve “’backwards” to its more treatable form. Clinicians could then use currently available drugs to treat the newly-non resistant infection.
More recently, scientists have used the term “steering” to talk about ways to manipulate populations of resistant organisms towards being treatable. Physicists and physicians have even used the rules of quantum physics to help think about ways to control the evolution of resistance.
But these applications aside, basic questions surrounding the forces that craft reversal are central in evolutionary theory. The answers are relevant not only for phenomena like antibiotic resistance, but also for other questions about cancer, resistance to butterfly toxins, and more. This even has implications for how we are thinking about genetic modification (GM): when we engineer a mutation into a crop for agricultural purposes, can we be sure that it won’t “un-evolve” that mutation “backwards” towards the non-GM crop?
Asked differently: can evolution do the moonwalk?
What is the moonwalk?
The moonwalk was made famous by Michael Jackson, but it was invented by Shalamar’s Jeffrey Daniel in 1982. We might take it for granted now, but when Daniel first pulled it off, it looked like a special effect. People thought that a string must have been pulling him because he seemed to be moving backwards just as seamlessly and smoothly as one can move forward.
So what does the moonwalk have to do with molecular evolution?
We can ask how evolution might move backwards—just like Daniel’s dance. Equipped with population genetic theory, we can examine the particular forces and conditions that facilitate that backwards movement.
Microbial moonwalking
I have previously published on reverse evolution in the context of antimalarial resistance. In that 2016 study, I found that the surprisingly large impact of certain compensatory mutations limits the ability of a population of resistant malaria parasites to become treatable again. Since then, I have remained curious about other ways to test Dollo’s law: what other places, models, and systems can we use to examine reverse evolution?
Luckily, other scientists I respect and admire had similar curiosities.
Dr. Pleuni Pennings is an evolutionary computational biologist who helped to pioneer the fusion of classical theoretical population genetics with very modern questions in HIV drug resistance. Her work had a very large impact on my career when I was learning population genetic approaches to thinking about disease evolution during my postdoctoral training. Although we’d already collaborated on several different educational programs and communication efforts, I had never had the pleasure of working with her on an actual science project until she approached me in 2019.
At that time, Dr. Pennings was in the process of establishing a collaboration with Dr. Ruth Hershberg, an eminent microbial evolution expert at Technion-Israel Institute of Technology. Dr. Hershberg had recently published a study that examined the results of experimental evolution in E. coli. Together, they were interested in exploring the conditions that facilitated reverse evolution in experimental populations of E.coli, and they brought me in on the question.
Over the next year or so, we discussed and designed several projects in the arena, but we decided that what we needed first was to use tools in theoretical and computational biology to ask some basic questions:
- How does mutation rate influence the probability of reverse evolution?
- How do the number and phenotypic effect of compensatory mutations influence the probability of reverse evolution?
These are two highly specific but very important variables to investigate for several reasons. For one, Dr. Hershberg had performed experimental evolution in background strains of E.coli that varied in their mutation rate. So we had the potential to compare computational results to experimental findings, which is an important point. Our collaboration provided the opportunity to use models and theory to explain experimental results. Plus, the questions surrounding the effects of mutations were compatible with prior work of mine about the nature and magnitude of compensatory mutations.
To ask these questions about reversion, here’s what we did:
- We created a “computerized world” where a population of bacteria had evolved resistance to a drug. In this situation, resistant variants carried a fitness cost in “drugless” environments, meaning resistant bacteria had high fitness in the presence of drug, but low fitness in the drugless environment.
- We then took the drug away (“drugless” environment) and asked whether the population of resistant bacteria would evolve “backwards” towards the wild type ancestor. We might expect this is because wild type had higher fitness than the resistant allele in the “drugless” environment.
- We specifically tuned two aspects of evolution in the “drugless” environment: mutation rate and the effect of compensatory mutation, meaning the degree to which mutations compensated for resistance.
The results were fascinating:
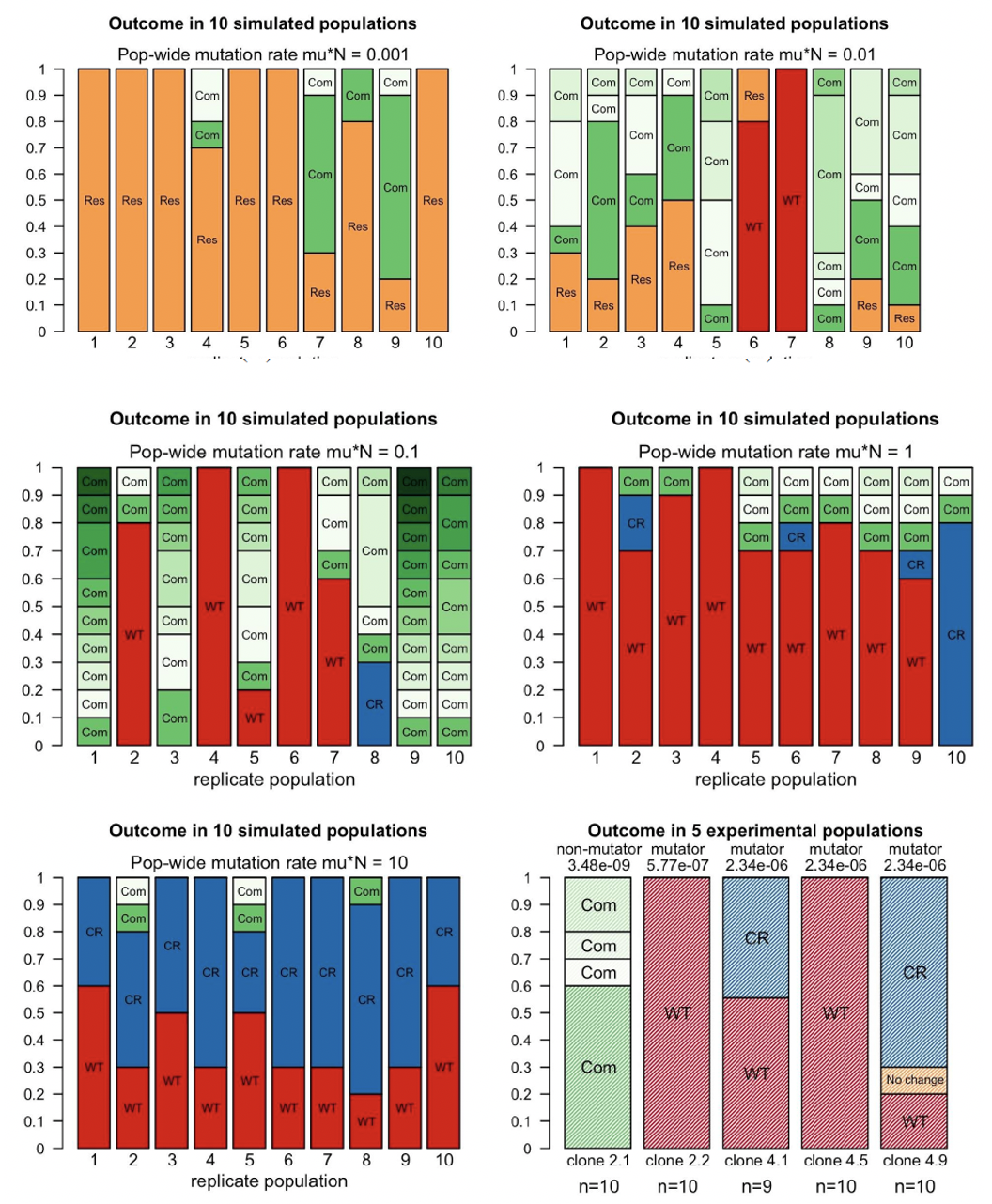
- The probability of reverting back to wild type was powerfully influenced by both the mutation rate and the effect of compensatory mutations. At very low mutation rates, compensatory mutations rose to fixation and no reversal to the wild type occurred. At very high mutation rates, compensated reversal emerged. Bacteria evolved two mutations: first the compensatory mutation, then the reversion to the wildtype. Intermediate mutation rates, however, displayed the proper conditions for reversal: the wild type mutation arose early enough that it could dominate a population (Figure 1).
- A lot of the findings really did boil down to how strong the compensatory effects were. For mutations that compensate for the cost of resistance: how many are there? Do they fully or partially compensate?
- Critically, the simulation results helped make sense of published experimental results. In these studies, reversal was observed when mutation rates were high. Our modeling result demonstrated that compensatory mutations in the experimental populations likely do not fully alleviate costs associated with resistance.
- This is a very big point: our simulations have diagnostic utility. From them, we can walk into experimental data and make sense of them.
In sum, the findings demonstrate how reverse evolution in the context of drug resistance is sensitive to population genetics particulars.
Why do we care?
For one, simply saying that reverse evolution is unlikely might not be wrong, but it is…incomplete. Population genetics can add color and rigor to this circumstance, so moving forward (no pun intended), all discussions of reverse evolution should be framed in terms of a particular population genetics setting.
Secondly, these results can inform modern conversations around how to steer the evolution of drug resistance. Perhaps we can rationally manipulate populations towards resistance or susceptibility using drugs of various kinds.
So, in the end, we can ask the bigger question:
Can evolution do the moonwalk?
The answer is “Yes, it can”—but the mutation rate and other properties of the context have to be just right.
And this is where the connection between the biology and the dance analogy emerge: you have to be a pretty creative person to come up with something like the moonwalk. But you can’t be too absurd. The innovative step that gave us the moonwalk was the product of an imagination with the right mutation effect size—one that imagines us moving backwards, against our intuition, but still right in rhythm.
Sure people might have been confused by the dance at first, but they will eventually realize that maybe their imaginations were too small and that there is no real forward and backward in dance—just like in molecular evolution.
CITATION
Reversion is most likely under high mutation supply, when compensatory mutations don’t fully restore fitness costs
Pleuni S Pennings, C Brandon Ogbunugafor, Ruth Hershberg
G3: Genes|Genomes|Genetics August 2022 jkac190; https://doi.org/10.1093/g3journal/jkac190
Guest posts are contributed by members of our community. The views expressed in guest posts are those of the author(s) and are not necessarily endorsed by the Genetics Society of America. If you'd like to write a guest post, e-mail jtreboschi@genetics-gsa.org.
View all posts by Guest Author »Read more in Science & Publishing
-

GENETICS welcomes new editor William Gilliland
A new associate editor is joining GENETICS. We’re excited to welcome William Gilliland to the editorial team. William Gilliland Associate Editor I have been a faculty at DePaul University in Chicago since 2009. I study female meiosis in Drosophila, and I’m most interested in topics like the movement of chromosomes through prometaphase I, the population…
-

New editors join GENETICS, G3 editorial boards
Several new editors are joining the GSA Journals. We’re excited to welcome Ricardo Zayas to the GENETICS editorial board under the Molecular Genetics of Development section, and on the G3: Genes|Genomes|Genetics board, we welcome Polly Campbell, Kevin Vogel, Joe Parker, and Ricardo Mallarino. Ricardo Zayas Associate Editor Ricardo Zayas is a Professor of Biology at…
-
Worms and Flies Provide Key Clues to Medical Mystery
This article is part of a series of posts outlining the history and impact of research in experimental organisms. The series is developed in collaboration with the GSA Public Communications and Engagement Committee. By the time Bertrand Might was six months old, it was clear something was amiss. His muscles weren’t developing normally; he was…