

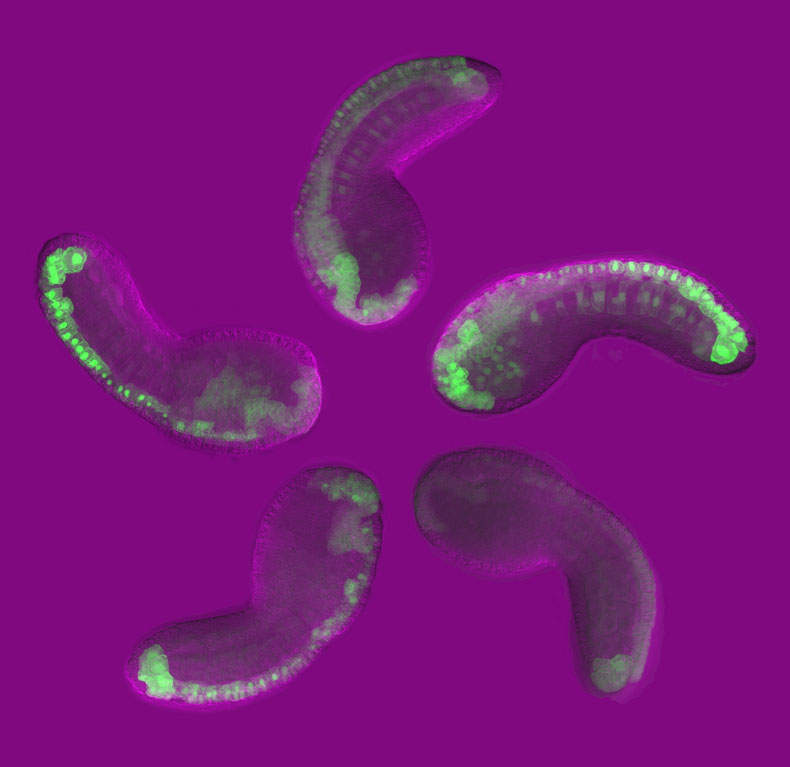
Today’s guest author is Emma Farley, a postdoctoral researcher in Mike Levine’s lab, which recently relocated to Princeton (formerly at University of California, Berkeley).
Sea squirts (Ciona intestanalis) are a classic system for the study of development. They were a favorite of early developmental biologists like Laurent Chabry, Ed Conklin, and Thomas Hunt Morgan. Over the course of 8 hours, you can watch a fertilized egg transform into a tadpole with a morphology indistinguishable from its vertebrate relatives! What’s more, the tadpole has only 2,500 cells. We can trace every single cell from the single celled fertilized egg to the fully formed tadpole because, unlike their vertebrate counterparts, sea squirts have defined cell lineages.
![Sea squirts. Image credit: Richard Ling [CC-BY-SA-2.0].](https://s43361.pcdn.co/wp-content/uploads/2015/11/8432568984_cca99cd469_z.jpg)
Sea squirts. Image credit: Richard Ling [CC-BY-SA-2.0]
The development of a sea squirt embryo follows a pattern similar to its vertebrate counterparts. This is not surprising when you consider that the sea squirt is a member of the ascidians, the sister group to the vertebrates. Sea squirts have been an ideal model organism for understanding the development and evolution of vertebrate-specific traits such as the origin of the two chambered heart and the origin of the vertebrate head. Another advantage of the Ciona is that its genome lacks the duplication seen in its vertebrate relatives, which makes it a simpler system for understanding development. For example, development of the Ciona heart uses the same genetic cues as mouse and humans, but is less complex, without the presence of multiple related proteins. These advantages all make Ciona an ideal system for studying the evolution of novel traits (e.g., neural crest and cranial placodes).
But there’s more to Ciona than evo-devo. As a postdoc in Mike Levine’s lab at UC Berkeley/Princeton, I was drawn to Ciona partly because we could introduce DNA constructs into thousands of embryos in a single hour-long experiment! Ciona—with its small number of cells, defined cell lineages, ease of imaging, and ease of introducing DNA—is an ideal system for high-throughput functional genomics, namely, how the genome encodes animal development.
![Sea squirts. Image credit: Prilfish [CC-BY-SA-2.0], via Wikimedia Commons](https://s43361.pcdn.co/wp-content/uploads/2015/11/930351131_55de272264_z.jpg)
Sea squirts. Image credit: Prilfish [CC-BY-SA-2.0], via Wikimedia Commons
It’s apparent that humans need their heart-related genes turned on in the heart rather than, say, in the skin. So how does the sequence of an enhancer ensure genes are turned on only in the right location? Enhancers contain hundreds of nucleotides and multiple binding sites for transcription factors. This means that even the smallest enhancer has 1030 potential sequence combinations and several hundred thousand possible “grammatical variants” (i.e., differences in the order and orientation of binding sites)! To understand how changes in enhancer sequence impact where and when specific genes are turned on, we need high-throughput functional assays in developing embryos. They allow for an assay in every tissue type and also ensure the same conditions the genome naturally experiences.
Importantly, several hundred thousand Ciona embryos can be electroporated in a single experiment. This makes Ciona an ideal system in which to carry out high-throughput functional genomic assays to understand how enhancers instruct development. As a postdoc, I conceived and developed Synthetic Enhancer Library-Seq (SEL-Seq), a method to create millions of enhancer variants, and then tested these variants for function in developing Ciona embryos.
We used this technique to investigate how an enhancer important for brain development turns on gene expression. We identified the sequences within the enhancer that are necessary and sufficient to turn on expression in the brain. Surprisingly, we found that these sequences possess a number of suboptimal properties (affinity of binding sites and spacing of binding sites) that make them poor for activating gene expression. When we improved these features, the enhancer went “haywire,” leading to expression in other tissues. We proposed that suboptimal features are used so that enhancers avoid accidentally turning genes on in the wrong part of the body.
The genome is filled with multiple enhancers that activate expression of the same gene in the same place. Why are so many enhancers responsible for the same job? Clusters of weak enhancers such as “super-enhancers” could circumvent a trade-off in expression specificity and activity, and generate robust but localized patterns of expression. Because switching on a gene in the wrong tissue can lead to disease, understanding the principles governing enhancer specificity should help us identify mistakes in our DNA that result in problems.
While Ciona is a model organism that is often dismissed as a boutique system, a closer look reveals that the sea squirt is far more powerful than its name! It is the perfect system for deciphering the instructions for development embedded in the genome.
{kind=link}

Emma Farley
Originally from Kent – the garden of England – Emma started in the Levine lab working on flies, but was quickly drawn to Ciona and the power of electroporation to decipher the regulatory code of the genome. Her specialty is transcriptional elongation and enhancer grammar. Ask her anything about enhancers – she enjoys discussing when, where, and why these strings of DNA sequence activate gene expression.
The views expressed in guest posts are those of the author and are not necessarily endorsed by the Genetics Society of America.
Guest posts are contributed by members of our community. The views expressed in guest posts are those of the author(s) and are not necessarily endorsed by the Genetics Society of America. If you'd like to write a guest post, e-mail jtreboschi@genetics-gsa.org.
View all posts by Guest Author »Read more in
-

Early Career Leadership Spotlight: Julio Molina Pineda
We’re taking time to get to know the members of the GSA’s Early Career Scientist Committees. Join us to learn more about our early career scientist advocates. Julio Molina Pineda Policy and Advocacy University of Arkansas Research Interest My research interests focus on using model organisms to genetically dissect complex traits related to human disease. My…
-

Early Career Leadership Spotlight: Peiwei Chen
We’re taking time to get to know the members of the GSA’s Early Career Scientist Committees. Join us to learn more about our early career scientist advocates. Peiwei Chen Accessibility Subcommittee California Institute of Technology Research Interest Far from a harmonious place, the genome is a battleground, where every bit of DNA fights for inheritance and…
-

#Dros23 GSA Poster Award winners
We are pleased to announce the GSA Poster Award winners from the 64th Annual Drosophila Research Conference! Undergraduate and graduate student members of the GSA were eligible for the awards, and a hard-working team of postdocs volunteered their time as judges. Congratulations to all! Undergraduate Students 1st Place: Sofia Karter Lopez, University of Toronto “Rab11 mediates E-cadherin recycling during…
-

Congratulations to the Fall 2022 DeLill Nasser Awardees!
GSA is pleased to announce the recipients of the DeLill Nasser Award for Professional Development in Genetics for Fall 2022! Given twice a year to graduate students and postdoctoral researchers, DeLill Nasser Awards support attendance at meetings and laboratory courses. The award is named in honor of DeLill Nasser, a long-time GSA supporter and National Science Foundation…
-

New editors join GENETICS, G3 editorial boards
Several new editors are joining the GSA Journals. We’re excited to welcome Ricardo Zayas to the GENETICS editorial board under the Molecular Genetics of Development section, and on the G3: Genes|Genomes|Genetics board, we welcome Polly Campbell, Kevin Vogel, Joe Parker, and Ricardo Mallarino. Ricardo Zayas Associate Editor Ricardo Zayas is a Professor of Biology at…
-
Worms and Flies Provide Key Clues to Medical Mystery
This article is part of a series of posts outlining the history and impact of research in experimental organisms. The series is developed in collaboration with the GSA Public Communications and Engagement Committee. By the time Bertrand Might was six months old, it was clear something was amiss. His muscles weren’t developing normally; he was…
-

Congratulations to the 2023 Early Career Leadership Program Cohort!
The Genetics Society of America (GSA) is excited to announce the latest cohort of student, postdoc, and early-career research leaders joining the Early Career Leadership Program. Participants receive training and mentoring while serving on committees charged with understanding the needs, interests, concerns, and challenges of early career scientist members of the GSA. As part of…
-

GSA LOCI: Local Outreach Community Initiatives @ GSA Conferences
Highlights: Local Outreach Community Initiatives (LOCI): The Genetics Society of America is committed to supporting the communities of the host cities of our conferences. This new year, we are excited to reconnect with our GSA community in meaningful ways within and beyond our existing programming. The GSA membership has created a caring and supportive environment…
-

New members of the GSA Board of Directors: 2023–2025
We are pleased to announce the election of five new leaders to the GSA Board of Directors: 2023 Vice President/2024 President Mariana Wolfner Distinguished Professor of Molecular Biology and Genetics and Stephen H. Weiss Presidential Fellow My research has focused on the genes and pathways that mediate sexual development and reproduction, primarily in Drosophila. From…
-

Lance David Miller: Lighting Your Own Fire by Finding the Right Resources
By Daniel J. Gironda In the Paths to Science Policy series, we talk to individuals who have a passion for science policy and are active in advocacy through their various roles and careers. The series aims to inform and guide early career scientists interested in science policy. This series is brought to you by the…
-

Graça Almeida-Porada: The Importance of Communication in a Technologically Advancing World
By Daniel J. Gironda In the Paths to Science Policy series, we talk to individuals who have a passion for science policy and are active in advocacy through their various roles and careers. The series aims to inform and guide early career scientists interested in science policy. This series is brought to you by the…